The Mother of All Placental Mammals Discovered: Identification of the first fossil Placental Mammal from the Cretaceous Period

Dr. Eugene Anklesloth
Every few years a team of researchers claims they've found a new ancestor of placental mammals, the member of clade Eutheria ancestral to all living placentals today. Unfortunately scientists have only tentatively identified a few species of Eutherian, much less one without an epipubic bone. The missing epibubic bone unites all living placental mammals, and is thought to be a loss necessitated by the need to carry offspring to term. The molecular clock results continue to estimate the divergence of all placental mammal groups to before the KTg extinction event that ended the dinosaurs, but still no true placental mammal emerges from the Cretaceous world.
This seems somewhat strange. The ancestors of marsupials and monotremes turn up regularly in the fossil beds, as do their extinct "sister groups" such as the Multituberculates and Allotheres. Some sister groups even survived into the Paleolithic Period, and others are extant today (Virginia Oppossum). We have plenty of fossils of all of these animals. Fossils of Tritylodontidae, a group which diverged from the ancestor of mammals over 230 million years ago, continue into the Late Cretaceous. Just like other mammals and their kin, archosaurs, dinosaurs, and birds provide a splendid fossil record all through the Jurrassic and Cretaceous periods.
But not the direct ancestor of placental mammals. Only a few tentative members of Jurassic and Cretaceous Eutherians have been identified. None of them have been shown to lack an epipubic bone. Attempts have been made to link certain Mesozoic "crown mammals" and extant placentals with features of the ankles, teeth, or skull. None of them have shown traits that mark them as a true placental mammal.
The first placental mammals do not appear in the fossil record until after 65 million years ago. This suggests a "ghost lineage" stretching back at least 160 million years, the time at which the ancestors of placentals and marsupials went their separate ways. Since monotremes and metatherians are considered divergent from a more "basal" or "unspecialized" lineage that ultimately led to placentals, it may indicate a ghost lineage that stretches back to 200 million years or more.
A ghost lineage is a procession of ancestral animal groups not evidenced in the fossil record but evident from the study of extant species or their DNA. Though many and long ghost lineages can and do exist, the lack of Eutherian fossils puzzles many, and the search for Jurassic and Cretaceous placental mammals continues.
It should be said that because of the ghost lineage, some researchers still hold to the idea that placental Eutherians only arose after the KTg event, despite the molecular evidence. The problem with this is threefold. After 66 million years ago, many placental animal groups would have had to cross thousands of miles of open ocean to get to where their fossils start showing up around 53 million years ago. The molecular dates line up perfectly with the fossil record for the divergence of monotremes and metatherians, and the latter two molecular dates should be more unreliable considering their much older age (200 million and 160 million respectively). Therefore the strict molecular dates for the divergence of placentals should be more accurate, and it always gives a date in the Cretaceous well before the Ktg event. (2) Third, such rapid evolutionary change (from 66 to 53 mya) from a shrew-like creature to whales, monkeys, and horned ungulates is unprecedented in more complete fossil records of other animal species. The differences between the earliest representatives of extant placentals are just too great to have happened in such a short time.
Regardless, the ghost lineage would still exist even under such a model, since Metatherians, Monotremes, Allotheres and others show clear divergence and specialization from a more "basal" or "generalized' lineage that is so far unrepresented in the Jurassic and Cretaceous.
But perhaps researchers have been searching in the wrong place.
There may yet be a few known species from the Late Cretaceous who almost fit the bill. Other than lack of hip girth and possession of an epipubic bone, Allotheres make pretty good candidates for the ancestors of placentals. Scientists think they diverged slightly before marsupials, but the classification of several species within the group is doubtful.
Some Gondwanatheres, tentatively placed into Allotheria, may be closer to placentals than marsupials. These herbivores reached among the largest size of any Cretaceous mammal. By the time of the KT event, they had become ubiquitous in several areas where the oldest placental mammal fossils have been found. Gondwanatheres survived the KTg extinction, and continued to thrive alongside placentals in some areas until as little at 17.5 million years ago.
But just like all other Allotheres, the Gondwanathere pelvis could not have handled carrying offspring to "term."
Of course, neither could the direct ancestor of the first placental mammal group. A series of mutations likely led to the enlarged womb and wider hips. Still, Gondwanatheres and Allotheres share features considered to be "derived" from the main lineage that led to placentals. This makes a Gondwanathere parent for placentals highly unlikely. Nevertheless, certain traits point to a close relationship with the most "primitive" form of placental mammal, Xenarthra.
"For several decades the affinities of the group were not clear, being first interpreted as early xenarthrans, or "toothless" mammals similar to the modern anteater. -Gondwanatheria, wikipedia
Recent studies have shown Xenarthra to be the most basal of all placentals, possibly at the root of the placental mammal family tree. This means they've retained many traits and genes which mutated and diversified in other placental groups since their last common ancestor. In at least one study, Xenarthra could not be ruled out as basal to placentals. This means some members of Xenarthra may not have changed significantly since 66 million to 120 million years ago, when the first superorder of mammals diverged from the common placental ancestor. (1)
Taken at face value, this could mean that a Gondwanathere begot Xenarthra, and Xenarthra begot all placental mammals. But there is more to link the two groups, and more to rule out such a notion.
Two "crown mammals" have been tentatively placed into Gondwanathere. Both bear even more resemblance to Xenarthra.
"Groeberia and Vintana both possess bizarre jugal flanges similar to those of xenarthrans like ground sloths." Gondwanatheria, wikipedia
Vintana


http://cdn.sci-news.com/images/enlarge/image_2256_2e-Vintana-sertichi.jpg
Giant Sloth Skull


http://www.angelfire.com/mi/dinosaurs/images/casts/sloth_eremotherium_v.jpg
Glyptodont Skull:


http://www.gastondesign.com/wp-content/uploads/clavipes-skull_fs.jpg
In fact the earliest known members of Xenarthra possess these structures, such as Glyptodont and Megatherium. The most basal form of Xenarthra, Dasypus, also has them, though reduced in proportion to the skull.
Such structures appear in the mammalia fossil record only 65.5 million years ago, and only appear in Groeberia, Vintana, and Xenarthra. So cheek flanges are a trait of primitive placental mammal and the only other mammal group that crossed the KT boundary with placental mammals—late, tentative gondwanatheres.
But one of these two specimens tells us that the trait likely goes back to 136 million years ago.
Vintana is a particularly important specimen. Being found on Madagascar with a date of 65.5 mya, it gives us a snap shot of what the ancestor of all mammals may have looked like during a period where fossils are scant. No earlier known mammal shares such a trait with a member of Eutheria. We don't know for certain if the species was "placental," because we've only found a skull.
However, Groeberia survived in S. America until only 28 mya, and left more fossils than Vintana. Since we can be sure Groberians were not placentals, we know the "cheek flanges" must have evolved before the first placental mammal arose. They must have been present in the Cretaceous ancestor of both placental mammals and Gondwanatheres(including Vintana), if not other groups. Otherwise glyptodonts and giant sloths and other Xenarthra wouldn't have them too.
So barring extreme coincidence and backward evolution, the first placental mammal sported "cheek flanges."
In fact, what might be remnants of cheek flanges can be seen on the skulls of some Afrotheres and Laurentians, such as the aardvark, dugong, manatee, and pangolin.
No Cretaceous mammaliaform displays these cheek-flanges, or even remnants/vestiges thereof. However, due to the date of Vintana we know the condition must have existed before the Cretaceous ended, and probably by 136 million years ago when India/Madagascar separated from the other continents.
So why can't we find the pre-KTg mammal who started it all? After KTg, fossils become rare for 10 million years or more. But directly before the KTg, we find many fossilized mammals. Why can't we find those cheek flanges on one of them?
I think we have. We found the ancestor long ago, but tradition has blinded our eyes.
But before I reveal my hypothesis, let's get a better picture of what our "crown placental" would look like.
We know the earliest Xenarthrans wore hard exoskeletons, from the giant sloth to Glyptodont. Whether they grew fur is unknown, though we assume they did since we have possible evidence of hair growth in mammaliaforms. But exoskeletons are made up of osteoderms. Osteoderms are comprised of bone and scutes, or "scales." We've got a pretty good idea scutes have evolved into hair more than once in the history of amniotes. Some species of proto-mammal (Therapsids) may have evolved hair from scutes independently of true mammals. Scientist debate whether the advanced scutes of certain ornithischian dinosaurs were proto-fur, fur, or feathers. Scutes may also become spikes, plates, horns, and whiskers. The pair of horns on ceratopsian dinosaurs are thought to be bone from osteoderms solidified to the skull. The origin of placental mammal horn bosses is simply a mystery which few scientists have even addressed. Until an explanation is forthcoming, we might assume that mammal horns evolved from osteoderms as well. But first we'd have to be sure an ancestor of horned mammals wore an osteoderm.
There's no good evidence that hair has ever turned back into scutes or scales. Perhaps it has, but we cannot confirm so at present. We can acknowledge that in order for hair to become scutes and reform into hair takes one step backward and another step forward in the mutation dance. Such processes require more "evolutionary effort" than a process in which scutes simply evolve into hair.
Because Xenarthrans are primitive mammals and have remained unchanged so long, we might guess the clade's common ancestor would have scutes as well as hair, maybe even an osteoderm. The original order of Xenarthra included pangolins and other animals ruled out by modern DNA testing. The group was based on certain "primitive" features, including homodonty or toothlessness and evidence of a present or former osteoderm. Common thought held the group evolved from an "armored" ancestor.
All species who remained in Xenarthrans after rigorous DNA tests exhibit a loss of "body armor" in their lineages. However, though studies have shown that the ancestor of all Xenarthra and the ancestor of all placental mammals may be the same or sister groups, no theories or studies have come forth whether this means the ancestor of all placentals possessed a shell of scute and bone (osteoderm).
However, as far back as the fossil record of Xenarthra goes, the fossils exhibit armor. And the most “primitive" xenarthrans haven't significantly changed in a minimum of 35- 45 million years.
This data combined suggests the ancestral placental may have had an osteoderm. Among placentals, only ungulates of clade Artiodactyla and the more primitive Xenarthrans possess true horns in the supraorbital position. A “shed” placental osteoderm provides an explanation for mammalian horns and remnants of osteoderms in former members of Xenarthra who have been reclassified, such as the pangolin.
What about teeth? Based on Xenarthra, what might "basal placental mammal" choppers look like?
Most if not all Xenarthra are homodonts, meaning they have undifferentiated teeth. Some don't have teeth at all. Most mammals, however, are heterodonts, meaning they have differentiated teeth. Reptiles are usually homodont, though there are many exceptions. Several dinosaur groups became as heterodont as mammals by the end of the Cretaceous. Pachycephalosauria had teeth like modern carnivores, and their close relative is aptly named Heterodontosaurus.
Heterodonty is agreed to be, to some degree, evidence of specialization to a niche environment. This is why tooth morphology is often used to identify relationships in fossil animals. Once a lineage's dentation becomes highly specialized, relationships between species can be established. Because our "flared cheek Gondwanatheres" had differentiated teeth similar to Allotheres and Xenarthra's teeth are homodont, we might suppose Gondwanathere divergent from a basal lineage. Madagascar and Chile make for pretty good niches.
We might conclude, therefore, that "basal placental" was a homodont. Basal can be seen as "unspecialized." If Xenarthra is our most unspecialized form of mammal, it follows that its morphology would be the least changed by time. The least steps to any mammalian order's overall tooth morphology is from a basal homodont state.
So it wouldn't be amiss to imagine our basal placental ancestor was a "homodont" too.
If we could find a Late Cretaceous mammaliaform with flared cheeks, scutes, xenarthic traits, xenarthic morphology, with evidence of an enlarged womb and birth canal, we'd be in business. We'd have our first known placental mammal.
But there's one more "trait" we might look for. So far, Vintana has been the closest thing to a "crown placental" we've identified. It's a crown mammal with cheek flanges at the right place at the right time. But it's hips are conveniently missing and it shares a few specialist traits with gondwanatheres.
It might behoove us to see what the scientists studying Vintana have to say about it:
"...revealed an utterly unique combination of features. Large eye sockets and nasal spaces, an inner ear designed for maintaining balance well and hearing high-pitched sounds, plant-grinding molars, and attachment sites for large incisors and powerful jaw muscles its estimated bite strength is twice that of the nutria, a similar-sized modern rodent suggest an active, agile herbivore with exceptionally keen senses for its time, able to eat tough material such as roots and large seeds. But some parts of its palate, nasal structure, and inner ear are 菟rimitive, having never been found in other true mammals but only their near-mammalian relatives such as tritylodontids"
Tritylodontids? But those aren't even quite mammaliaforms. They arose 221 million years ago, just before marsupials split off from the ancestor of marsupials and placentals. Only a few survived the extinction Triassic Extinction event to re-diversify. They may or may not have directly given rise to the mammals. Their resemblance to Vintana might surprise at first, but it actually makes sense.
Monotremes and Marsupials are specialists, divergent from the main lineage that ultimately led to placentals. And Tritylodontids existed until 113 million years ago, near the molecular "strict date" for the direct ancestor of all extant placental mammals. If not the lack of a few mammalian traits, such as the mammalian jaw, they might make another good candidate for our "basal crown placental."
The connection with Vintana might make Tritylodontids a sibling group to the "ghost lineage" from which marsupials and monotremes diverged over 150 million years ago—the elusive lineage Eutheria, which led to placental mammals. Therefore it's to be expected this ghost lineage would retain a few Tritylodontid traits not found in divergent groups.
But Eutheria and placental Mammalia have been ghost lineages for a long time. We have fossils from many different groups that arose from our direct lineage, but not our direct lineage itself. We have plenty of fossils of Allotheres, Multituberculates, Marsupials, Monotremes, and several more groups which split off from this basal lineage. But we don't have the basal lineage. We don't have the ancestors from which all of these groups arose, the lineage that led to the first placental mammals.
The Molecular clocks report dates for placental mammals diverging during the Aptian Age, when the strait between Africa and Laurasia widened to an ocean and the continents began to move away from one another. The period is more than 50 million years before the first true placental mammal appears in the fossil record. Why not even one measly fossil? One hip bone that demonstrates the ability to give live birth to largish children? Without the epipubic bone?
We might find a clue in the Tritylodontid traits of Vintana.
We might look for the same reptilian traits in our Late Cretaceous "basal placental mammal." Our "ghost lineage" most likely retained them after Gondwanatheres split off, and even retained them until becoming Xenarthra (or long after).
And since Xenarthra and Vintana had a keen sense of smell and bulbous eyes we might look for those traits in our first placental mammal as well.
If we could find a mammaliaform with flared cheeks, scutes, xenarthic traits, xenarthic morphology, body-armor, wide hips with access for a womb and birth canal, a good sense of smell, hearing, large eyes, an osteoderm and a few primitive skull traits we'd be looking good.
Just the kind of animal for Tritylodontids to diverge from. Or monotremes, marsupials, and Gondwantheres. Just the kind of thing to diverge into all the varied groups of placental mammal.
And I bet an animal that retained such traits might be able to survive an apocalypse or two, followed by a few years of total darkness. A walking tank with nutcracker jaws and the ability to swim, climb, crawl, and perhaps roll up in a ball. An omnivore with a long tongue not too picky whether it's bugs or plants for dinner. Resistance to infection and disease might not hurt either...and animals covered with scutes certainly have it.
Pangolin genomes and the evolution of mammalian scales and immunity
The lineage which led to extant placental mammals survived more at least six apocalypses while retaining a similar morphology. At least two of these global disasters ended the majority of terrestrial life. This ghost lineage retained basal traits while whole orders of animal species arose, diversified, and went extinct over and over again, from tritylodontids to allotheres to dinosaurs. Our Triassic/Jurassic/Cretaceous ancestor must have been damn tough to have survived when nearly all others failed. And Vintana's anatomy tells us the lineage retained certain generalized traits for over 100 million years.
Is there an animal in the Maastrichtian(Latest Cretaceous) which might fit the bill? Of course it would have to be a synapsid. All mammals are synapsids. That's one of the main ways we tell them from reptiles when we find a new fossil.
Could there be a species whose skeleton morphology, bone fusions, and dentition more closely matches Xenarthra than any other from the period? Is there a species with skull and cheek flanges similar to these groups we've shown to be related? A synapsid with all of the traits we've been looking for?
I'm sure of it.
Vintana may have even been a juvenile of the species or an example of "insular dwarfism." Because India/Madagascar spit from Gondwanaland 135 million years ago, we can assume Vintana split off from a mammalian ancestor with cheek flanges prior to that date. Only one group of animals from the Cretaceous sported any kind of cheek flanges.
Ornithischians. Specifically Stegosaurs, Ankylosaurs, and Ceratopsians. Many species within these groups reverted to a synapsid state during the Aptian Age. The evidence points to a synapsid ancestor for all of them, who lived at the same time as the Tritylodontids during the Late Jurassic. And Late Cretaceous members of this group and their closest ancestors show clear signs of both an enlarged hip cavity, broadened hips, and a canal in the vertebrae. These may be signs of an evolutionary change to holding infants to term.
As amazing as this may sound, Ornithiscians were mammals, and soon after 156 million years ago several species of Ornithiscians became placental mammals.
Ankylosaurs and their closest relatives show a clear precursor to the Gondwanathere/Xenarthran condition in their cheekbones. But flanges make but the weakest support in a mass of tumultuous proof. I will try to relate much of the mounting evidence in the following few pages.
Placental mammals evolved from ankylosaurus.
Contrary to common belief, genus ankylosaurus was made up of cynodonts, and the traits which separate it from Stegosaurus are analogous to several found in placental mammals.
“The loss of the antorbital and supratemporal fenestrae occurs by expansion of the adjacent bones closing the openings (a dimple may mark antorbital fenestra in some ankylosaurs.)”
““The snout of Ankylosaurus was arched and truncated at the front, and the nostrils were elliptical, and they were directed downward and outward, unlike in all other known ankylosaurids where they faced obliquely forward or upward. Additionally, the nostrils were not visible from the front, because the sinuseswere expanded to the sides of the premaxilla bone, to a larger extent than seen in other ankylosaurs. Large loreal caputegulae, strap-like, side osteoderms of the snout, completely roofed the enlarged opening of the nostrils. The nostrils also had an intranarial septum which separated the nasal passage from the sinus. Each side of the snout had five sinuses, four of which expanded into the maxilla bone. The nasal cavities or chambers of Ankylosaurus were elongated and separated by a septum at the midline, which divided the inside of the snout into two mirrored halves. The septum had two openings, including the chonea (internal nostrils).” -Ankylosaurus, Wikipedia Nov.7 2017
“Closure of antorbital and upper temporal fenestra, ossification and fusion of keeled plate onto side of lower jaw, fusion of first tasil vertebrae sacral vertebrae and ilium, rotation of ilium to to form flaring blades, closure of hip joint, development of dorsal shield of symmetrically placed bony plates and spines.” Dinosaurs, A Concise Natural History Cambridge University press 2016
Ankylosaurus possessed all of the traits listed above, even clear evidence of a womb and adaptions to pregnancy. Like Multituberculates, Allotheres, Gondwanatheres, early Xenarthrans and other basal placentals the snout had become elongated and olfactory adaptation is evident. Even the skull position and shape shows a transition toward placental mammals.
Scientists used to think the hip cavity and canal held an extra brain. While the hips of ankylosaurus have not been preserved, we know that stegosaur and its closest relatives had this feature. And the closest of all to stegosaurus is ankylosaurus.
“Soon after describing Stegosaurus, Marsh noted a large canal in the hip region of the spinal cord, which could have accommodated a structure up to 20 times larger than the famously small brain. This has led to the influential idea that dinosaurs like Stegosaurus had a "second brain" in the tail, which may have been responsible for controlling reflexes in the rear portion of the body. This "brain" might have given a Stegosaurus a temporary boost when it was under threat from predators.[12] More recently discussed, this space (also found in sauropods) may have been the location of a glycogen body, a structure in living birds whose function is not definitely known, but which is postulated to facilitate the supply of glycogen to the animal's nervous system.” Stegosaurus, wikipedia
Thyreophora contains mainly stegosaurs and ankylosaurs, and appears in our fossil record 200 million years ago.The few Jurassic members outside Stegosauria and Ankylosauria may or may not be misclassified. Interestingly, several of them are small specialists with decidedly mammalian teeth.
Earlier members of Ankylosauria and Thyreophora were non-placental mammals. The "brain" and "canal" appears after 156 million years ago in stegosaurus and certain relatives. The more placental Ankylosauridae family appears around 136 million years ago, soon after stegosaur. Stegosaur and members of the Ankylosauridae family are considered by many to be more related to one another than either are to other Ankylosauria and other Thyreophora species.
But there is evidence of a tendency toward larger wombs and mammalian traits in all ornithischians:
Ornithischians had paired premaxillary bones that were toothless and roughened at the tip of the snout (presumably due to the attachment of a keratinous beak).[2]
- Ornithischians developed a narrow "eyebrow", or palpebral bone, across the outside of the eye socket.[2]
- Ornithischians had reduced, or even closed-off, antorbital fenestrae (the fenestra in front of the eye socket).[2]
- Ornithischian jaw joints were lowered below the level of the teeth, bringing the teeth into simultaneous occlusion.[2]
- Ornithischians had "leaf-shaped" cheek teeth.[2]
- Ornithischian backbones were stiffened near the pelvis by the ossification of tendons above the sacrum. Additionally, ornithischians had at least five sacral vertebrae attaching to the pelvis.[]Ornithischians, wikipedia
Sometime before 88 mya, a species of Ankylosauridae lost its supraorbital fenestae and became genus Ankylosaurus, the direct ancestor of all placental mammals.
We can assume ankylosaurus would have shared the features of stegosaurs and other ornithiscians, but no feet or hips of ankylosaurus have been preserved to us in the fossil record.
Or have they?
Ankylosaurus is often found with the bones of Multituberculata, a sister group of placental mammals and marsupials somewhat synonymous to Alllotheres. Yep, the family of Vintana and gondwanatheres.
What's more, Multituberculata/Allotheres share tooth traits with the earliest whales, as well as extant whales. These are called tubercles. They make certain teeth a bit leaf-shaped, so to speak. They also possess a tooth called the Plagiaulacoid. It's present in the most primitive marsupials and the earliest proto-primates. It's found in other basal species of other placental mammal groups. Why wikipedia says it's convergent evolution is beyond me. Everyone knows all of those groups split from a common ancestor. There's nothing to say that ancestor didn't have a Plagiaulacoid.
In fact the ancestor did—it's name is Ankylosaur.
Of course scientists call the tooth by a different name. Ornithischians simply have an extra pair of teeth—just like all early mammals.
Interestingly, ankylosaurs and multituberculates are often found in the same dig site. Often, they only find the shells and skulls of the ankylosaurs, and the limbs of the multituberculates. Perhaps it's just another coincidence?
One day, we may find that ankylosaur and multituburculate remains from the same deposits to be from the same creature, or the same species. Future investigation may show that juveniles, adults, males, and females of the same species have been misidentified as separate species. This happens all the time, and we've discovered such errors often even within the last five years. Incidentally, one of those misidentifications involves a close relative of Ankylosaurs—Ceraptops, a group we'll discuss in a moment.
All mammaliaformes from the Jurassic and Cretaceous periods split off from the basal ankylosaur form(Thyreophora). Loss of temporal fenestra seems to be a result of specialization in these groups. Downsizing and a change of diet, somewhat akin to “insular dwarfism,” seems to accompany the loss of supraorbital fenestra. A smaller size than ancestral lineages is evident in ornithiscian synapids like certain ankylosaurus species and it's closest relatives, Pachycephalosaurs and Ornithopods. Ceratopsians, the fourth “sister species” of the Late Cretaceous Ornithischians, lost antoribital fenestra at a similar date. Several “dwarf species” appear just before the Ktg event, along with other mammalian traits. Sounds like a time when niches formed and animals changed their diets.
These four related animal groups lost fenestra and downsized at roughly the same time during the Aptian Age, when the atmosphere began to change rapidly and sea levels rose. During this time, the true dinosaurs grew to their largest size. Oxygen levels dropped to 15% lower than today and grass began to grow for the first time ever.
Supraorbital fenestra serve as attachment points for jaw muscles. Ornithischian supraorbital fenstrae never grew as large as archosaurs or Saurischia (true dinosaurs). Unlike most other diapsids, ornithischian temporal fenestrae were positioned more to the side of the skull. Since ornithiscians became synapsids independently of one another at the end of the Cretaceous, perhaps their ancestor had become a "diapsid" in response to a change of diet and increased jaw size. Maybe their reversion to a synapsid state marked another dietary switch during a rapidly changing time period.
Could the ornithischians have evolved from Therapsids, just as mammaliaformes are thought to have done? If so, can we find a transitional species between the late Triassic Tritylodontids and the early Jurassic Ornithiscians?
Aetosaurs were far more mammal-like than the therapsids of their time, but have been overlooked because they are classified as archosaur diapsids. Like ornithiscians and unlike other archosaurs, their temporal fenestrae sit on the sides of their skulls and their supraorbital fenestrae are small.
“New synapomorphies included temporal fenestrae, or holes, that opened on the side of the skull rather than the top...”
Aetosaur, Wikipedia
They are one of the few archosaur groups thought to have been herbivorous, and evolved to chew leaves and dig roots with pulling teeth. They were also insectivores. Their placement in archosauria is largely based on their classification as diapsids and their osteoderms; among “terrestrial” amniotes, only crocodylamorphs and turtles possessed body armor in the Triassic.
“Although aetosaurs are known exclusively from the Late Triassic, their currently accepted position in archosaur phylogeny indicates that they originated from more basal pseudosuchian archosaurs in the Early or Middle Triassic. Given that aetosaurs are highly specialized with many anatomical features not seen in other pseudosuchians, the group's evolutionary origins are poorly understood. “ Aetosaur, Wikipedia
We may even have a “transitional fossil” from synapsid to Aetosaur:
“In 2012 a third "stem aetosaur" was described from the Middle Triassic Manda Beds of Tanzania. It differs from other Middle Triassic pseudosuchians in having a long skull, a small antorbital fenestra that fits into a large antorbital fossa in front of the eye socket, sharp and curved teeth, and osteoderms covering much of its body. Like aetosaurs and Revueltosaurus, it has a maxilla that fits into the jugal.” Aetosaur, Wikipedia
Aetosaurs share morphology and many traits with armadillos, pangolins, and ankylosaurs. Just the thing to survive the Triassic-Jurrassic extinction event, and pass the torch on to Thyreophora. They appear at the same time as Tritylodontids in our fossil record, and end at the extinction boundary point. A new clade of Tritylodontids arise shortly after the extinction boundary at the same time as Thyreophora.
Amazingly, the skull shape from Aetosaur to Armadillo has barely changed in 200 million years. It's just gotten smaller and closed the fenestrae.
Aetosaur


Armadillo


https://farm3.static.flickr.com/2573/3945029267_8a67663a98_b.jpg
But whether Therapsids and Thyreophora diverged from Aetosaurs or Aetosaurs diverged from Therapsids, our facts continue to line up.
It would be easy to say genus Ankylosaurus diversified into all known groups of placental mammals sometime between 136 million and 66 million years ago. We could then uphold the idea that all placentals evolved from a common synapsid ancestor. However, that's not exactly how it happened. Ankylosaurus may have already become Xenarthra by the Late cretaceous. An earlier member of Thyreophora, such as an earlier ankylosaur or stegosaur, is more likely the ancestor we're looking for.
Prior to 117 million years ago, land bridges existed between Laurasia (Eurasia and N. America) and Africa. Only a short channel separated Africa from S. America. Between 117 million and 93 million years ago, the Aptian extinction event and two other volcanic apocalypses bottlenecked marine life, rose sea levels, and mark an increase in plate tectonic activity. After 93 million years ago, thousands of miles separated the continents and most remained isolated until well after 50 million years ago.
Though ankylosaurs are found on almost every continent during the Late Cretaceous, they are not found in Africa. In broken Laurasia and Antartica they persist until the time of the KT event.
But there are several other Late Cretaceous ornithiscian groups who had begun to lose fenestrae by the time of the KTg, and they all happen to be most closely related to Ankylosaurus. Further, they are at the right place at the right time, extant until 66 million years ago in the same places where mammals with similar morphologies appear after the Ktg event. Therefore, it's more likely that becoming synapsid was a trend among a group of closely related species. As we've seen, they were likely reverting to an original ancestral state in response to a change of diet and a process similar to “insular dwarfism.”
Beginning 200 million years ago, groups of mammal split off from Thyreophora(Aetosaur?) and became various Therapsid groups, including monotremes, metatherians, multituberculates, marsupials, and gondwanatheres. Divergence often entailed loss of fenestra and downsizing. The earliest to split off retained the “duck bill” of Aetosaur and Stegosaur for a time, as well as the smallish Aetosaur cheek flanges . The last to split off share enlarged cheek flanges, like Vintana, Goeberia, Ankylosaurs, and Ceratopsians. The large jaws of “dwarf species” and juveniles are evidence of evolution to a smaller skull while retaining the same eating habits.
Cerapoda split off from Ankylosaurus prior to 93 mya when the sea between Laurasia and Africa widened, cutting off land bridges. Cerapods diverged into Ceratops, Pachycephalosauria, and Ornithopoda .
One of many paths which led me to fascinating realizations about Earth's past presented itself in the ridiculous coincidence of horns. Their are only two groups of animal in over 350 million years of terrestrial amniote evolution who display a diversity of true horn bossess in the supraorbital position. Both of these groups are hooved herbivores with the same number of toes. They both evolved in Laurasia when Laurasia was separated from the other continents by several thousand miles. But these two groups are not thought to be related.
According to the strict molecular date Artiodactyla, horned/hooved ungulates, diverged from the original placental mammal 75 million to 114 million years ago. This is the time of the first two Late Cretaceous extinction events, when oxygen levels dipped to 15% lower than today and many species, especially marine, perished. Sea levels rose and the continets moved farther away from each other. The straits between Africa and S. America widened, as did the channel between Laurasia and Africa. All land bridges were flooded by 93 mya.
Exactly during this time period, the Ceratopsians appear in Laurasia.
The current model suggests that placental mammals and ceratops evolved horn bosses, hooves, a like number of toes, ruminating abilities, and a herding lifestyle at the exact same time in Laurasia.
An alternative to this is that the ancestral Artiodactyla swam thousands of mile to Laurasia from Africa and sprouted horn bosses and the above traits just 35 million years after the Ceratopsians.
Either way it's quite a coincidence. But maybe we can use Occam's Razor to divulge a more likely scenario. One with fewer assumptions and coincidences.
Ceratops evolved into Artiodactyla. This is why the only time in 350 million years a group of animals sprouted horn bosses in a multitude of styles was in Laurasia between 100 million and 50 million years ago. Laurasia did not cause both dinosaurs and mammals to suddenly evolve horns as mainstream science claims. Ceratops and horned ungulates were one in the same, and the trait must have become useful in the basal forefather of Ceratops/Artiodactyla. By the time of the KT event, ceratopsians were losing fenestra as the fossil record shows. By 53 mya, they'd lost their supratemporal fenestra as well.
HORNS
To be sure, a few other species of animal sport pairs of horns in the supraorbital region. These happen to be armadillos and Ankylosaurs. In both groups, the horns disappeared in most species and never developed into a variety of bosses.
And ankylosaurs and ceratopsians are sibling species. Armadillos are the most basal form of placental mammal. The possibility that Artiodactyla evolved directly from a species homologous to Xenarthra cannot be ruled out. The last common ancestor of all Xenarthrans may be the the last common ancestor of all placental mammals.
And Ankylosaurus may be analogous to superorder Xenarthra. No two misclassified animals have ever been more alike, in both superficial resemblance and highly scientific anatomical comparison. A species of Ankylosaurus became Xenartha shortly after the KT event some 66 million years ago, and the evidence is undeniable.
Some scientists have proposed ceratopsians sported beaks because of the missing portion of their lip. Cows and giraffes have remnants of this adaptation:
"Like domestic cattle, instead of upper teeth, giraffes have ・dental pads.” In actuality, the dental pads in giraffes are less evolved than their bovine brethren. Instead of thick wide pads, giraffe’s are hard and narrow, making it easier to crush their hard food. “
(3)
They also share a sort of gum covering like that of Allotheres.
“Since they mostly consume thorny acacia trees, their dental structure had to evolve to handle the sharp twigs. Giraffe tongues are covered in a thick papillae and gelatinous saliva, so they're not able to feel the sharp thorns. “
(3)
Scientists have proposed a beak instead of gum coverings for the ornithiscians, but the missing portion of jaw is the same. There is no actual evidence that any keratin beak ever existed.
One species of Ceratops even began to chew like an herbivorous mammal just before the Ktg. The distinction of chewing styles and dental morphology and wearing patterns has often been used to distinguish dinosaurs and mammals. This knowledge will be important for our conclusion:
Chew on this! Mammal-like mastication for the dinosaur Leptoceratops.
(4)
(4)
Ornithopoda evolved into Macraucheniidae. Most members of these two groups have the same number of toes. This is why Macraucheniidae bears a vestige of the Ornithopoda ornamental head outgrowth in the shape of its skull. Several species of Ornithopoda had already lost this outgrowth by the time of the Ktg. In fact, some hadrosaurs had already lost their supraorbital and antorbital fenestra and downsized by the end of the Cretaceous.(5)
“In contrast, most ornithischian dinosaurs reduce and even close their antorbital fenestrae[3] such as in hadrosaurs and the dinosaur genus Protoceratops.“ornithischian, wikipedia
“Maiasaura peeblesorum has a number of primitive characteristics, including an elongated facial region, restricted nares, deflected premaxillae, and an apparent remnant of the supraorbital fenestra “
(6)
(6)
Pachycephalosauria were also synapsids, just like Ankylosaurus and some Hadrosaurs. They and their closest relatives evolved into Euarchontoglires (or proto-primates within the group, such as adapids). The heterodont teeth of Pachycephalosauria can be mistaken for those of mammals. Being bipedal, Pachycephalosauria can also be mistaken for Sauropod dinosaurs.
“The characteristic skull of pachycephalosaurs is a result of the fusion and thickening of the frontals and parietals, accompanied by the closing of the supratemporal fenestra. In some species this takes the form of a raised dome; in others, the skull is flat or wedge-shaped. While the flat-headed pachycephalosaurs are traditionally regarded as distinct species or even families, they may represent juveniles of dome-headed adults.[2] All display highly ornamented jugals, squamosals, and postorbitals in the form of blunt horns and nodes. Many species are only known from skull fragments, and a complete pachycephalosaur skeleton is yet to be found." Pachycephalosaurs, wikipedia
squaキmoキsal
skwāˈmōsəl/
noun
ZOOLOGY
- the squamous portion of the temporal bone, especially when this forms a separate bone that, in mammals, articulates with the lower jaw.
- In many mammals, including humans, it fuses with the periotic bone and the auditory bulla to form the temporal bone, then referred to as the squama temporalis. (Wikipedia-squamosal)
“The postorbital is one of the bones in vertebrate skulls which forms a portion of the dermal skull roof and, sometimes, a ring about the orbit. Generally, it is located behind the postfrontal and posteriorly to th orbital fenestra. In some vertebrates, the postorbital is fused with the postfrontal to create a postorbitofrontal. “ postorbital, wikipedia
You can get a great sense of how placental mammal skull evolved by looking at the skulls of the youngest ankylosaurs (66 mya), followed by those of the youngest pachycephalosaurs, followed by those of the oldest Euarchontoglires (primates and rodents. 55+/- mya).
Ankylosaurid


Basal pachycephalosaurs



{kind=link}
{kind=link}
{kind=link}
https://i.pinimg.com/236x/a6/71/f8/a671f80097f0e3ff42a51d15a7d189dd--extinct-skull.jpg
Late pachycephalosaurs


{kind=link}
Euarchontoglires Adapids (proto-primates)


http://l7.alamy.com/zooms/aefc89dab6344b27b91030b34176b294/adapid-fossil-skull-adapids-are-lemur-like-primates-d3fdmb.jpg


https://i.pinimg.com/564x/0b/a5/9b/0ba59b25a9110c7e7d50e97406964a60--primates-mammals.jpg


http://bio.sunyorange.edu/updated2/pl%20new/Ch.%2069--Primates.Prosimians_files/23%20lemur.jpg
By the time of the youngest and/or smallest pachycephalosaurs, the blunt horns and nodes are mostly or completely gone. The beginnings of the zygomatic arch can be seen in a few specimens.
We may have even found another stage of evolution in the skull of Zhul, a new species of Ankylosaur:
“The caputegulae on the prefrontals, the frontoparietals and the middle supraorbitals are pyramid-shaped, in contrast to the conical caputegulae with Nodocephalosaurus and Talarurus. The squamosal horn protrudes to behind the rear edge of the skull roof, just as with Scolosaurus but different from Anodontosaurus, Euoplocephalus or Ziapelta. The caputegulae behind the eye socket are small and sparsely distributed, again like Scolosaurusbut differing from Anodontosaurus, Euoplocephalus or Ziapelta. The osteoderms on the handle of the tail club are relatively larger and more pointed than those of Asian ankylosaurines of the Nemegt Formation of Mongolia.” Zuul, Wikipedia
After the KTg event, surviving subspecies of pachycephalosaur likely rejoined with neighboring sibling subspecies in the process of assimilation. This is exactly what John Hawks and other scientists claim happened to our own species in Africa 200,000 years ago. In the barren conditions following the KTg, closely related subspecies mixed to survive. They were forced to economize their morphologies. The process may be likened to insular dwarfism. Many adaptations were needed to survive a period of recovery.
And so hybrid species of Dwarf pachycephalosaur became Euarchontoglires and their close relatives.
“Euarchontoglires probably split from the Laurasiatheria sister group about 85 to 95 million years ago, during the Cretaceous, and developed in the Laurasian island group that would later become Europe. This hypothesis is supported by molecular evidence; so far, the earliest known fossils date to the early Paleocene.[3] The clade of Euarchontoglires and Laurasiatheria is recognized as Boreoeutheria.[citation needed] Both Euarchontoglires and diprotodont marsupials are documented to possess a vermiform appendix, although this evolved as a result of convergent evolution.[4]“ Euarchontoglires, wikipedia
There's really no reason venom has to be convergent evolution. They provide no impetus for the similar development of venom, and they have not identified fossil specimens of the direct common ancestor of the two mammalian groups. A few have been proposed but will likely be proven divergent from the basal lineage. Until scientists seriously consider Ankylosaurus and pachycephalosaurs, that is.
Perhaps the ghost lineage they're looking for (Eutheria) had a bit of venom too. Most placentals simply lost it. It cost too much "evolutionary effort" to keep.
The periods after the Aptian Extinction Event and the KT event seem to have been accompanied by economizing. A transition to synapsy seems to have occurred over multiple lineages within Thyreophora, including monotremes, marsupials, ornithopods, pachycephalosaur, and late ankylosaurus during the filling of niches in the Jurassic and Cretaceous.
Monotremes, marsupials, and placentals have only remnants of temporal fenestra. This is a rarity in the fossil record before the Ktg event. Because no direct ancestor of mammals in the Cretaceous displays a closed fenestra and zygomatic arch like extant mammals, it can be seen as a result of the same process. Closing of both pairs of supraorbital fenestra occurred simultaneously among many related species at the end of the Cretaceous, possibly as a response to downsizing, climate, change, and new herbivorous opportunities.
Pachycephalosaurs also possess the potential for a birth canal, unlike all other crown mammals of the Late Cretaceous(with the exception of other ornithischians).
“They had a bulky torso with an expanded gut cavity and broad hips, short forelimbs, long legs, a short, thick neck, and a heavy tail “ Wikipedia's Pachycephalosaurs
No ornithiscians had epipubic bones.
The biggest difference between Pachycephalosaurs and early Euarchontoglires is that the lower temporal fenestra had expanded to complete the zygomatic arch in the latter. The second biggest difference is that Euarchontoglires had become arboreal, and hence had longer arm spans. There is more similarity between the two species than there is between early Euarchontoglires and any pre-KT mammal.
“There is an academic debate on the time the first primates appeared. One of the earliest probable primate fossils is the problematic Altiatlasius koulchii, perhaps an Omomyid, but perhaps a non-Primate Plesiadapiform, which lived in Morocco, during the Paleocene, around 60 Ma.[1] However, other studies, including molecular clock studies, have estimated the origin of the primate branch to have been in the mid-Cretaceous period, around 85 Ma, that is to say, in the time previous to the extinction of dinosaurs and the successful mammal radiation. Nevertheless, there seems to be a consensus about the monophyletic origin of the order, although the evidence is not clear.” -Wikipedia
Prediction: more fossils will be found between 53 million years ago and 66 million years ago. It will become increasingly difficult to tell the difference between early Euarchontoglires and late Pachycephalosauria.
Now monkeys don't need to swim thousands of miles to S. America in groups of 30 or more, as the old model suggests. Pachycephalosauria is last found in Asia 66 million years ago and is also found in North America at late dates. Euarchontoglires shows up in Asia around 53 million years ago, when fossils begin to show up for all other major placental mammal orders.
53 million years is the date at which life had fully recovered from the KT event, and animals became prolific enough to leave an abundance of fossils.
Pachycephalosauria/ Euarchontoglires island hopped to S. America around 20 million years ago, just before the land bridge between Americas fully formed. And thats when we find the first primate fossils in S. America.
I think they island hopped to Africa from Eurasia around 85 thousand years ago, when there was still a chance to do so. We'll eventually find a fossil or two. We may already have, and then misclassified it as something else. The smallest animals leave the least and most impartial remains.
Pachycephalosauria were among the first to specialize and downsize, hence an earlier loss of the cheek flanges and supraorbital fenestra.
Perhaps the most convincing argument is the so-called convergent evolution of Mosasaurs and Cetaceans.
The mainsteam evolutionary model requires a ghost lineage for mosasaurs that stretches back longer than these dinosaurs, possibly the same ghost lineage from which varanid lizards arose. Mosasaurs are thought to share a relationship with veranids and snakes, with a possible synodont ancestor in the Carboniferous or Triassic. So why did two groups of land animal, completely unrelated, evolve the same fish-like morphology in 350+ million years of amniote evolution? Artiodactyla, the clade of Cetaceans, diverged sometime after 100 mya ago. Mosasaurs appear 92 million years ago, roughly the same time...seemingly out of nowhere. Except for number of temporal fenestrae, their morphologies are the same as early whales, down to the enlarged abdomens rare in actual fish. Even the number of vertebrae is the same. Even the placement of the nostrils:
“I noticed two ridges on the top of the skull that indicate this fossil was geologically younger than others,” says Konishi. “That means this specimen was a more evolved mosasaur and I think the ridges indicate the animal’s breathing holes were set further back on the skull than earlier species.” The same pattern can be seen in modern animals; early mosasaurs can be compared to seals and walruses, which still have nostrils close to the front of their skulls. Later ones would have been more like whales and dolphins, with blowholes getting further toward the top of the skull. “ Takuya Konishi
Even the undifferentiated teeth of modern toothed whales, a decisively un-mammal-like trait, is extremely similar to those of the mosasaurs. Artiodactyl, the order of Cetaceans, diverged from placentals at the same time mosasaurs appear or slightly before, according to the “strict” molecular clock. But the first cetacean appears as a fossil 52 mya, almost simultaneously with the first appearance of a basal member of their order, artiodactyl. Yet only once in 400+ million years did such animals with the morphology of mosasaur and cetacean arise on the planet. These two animals arose from reptiles and mammals at roughly the same time or within 36 million years of each other at the longes stretch(Himalayacetus). Neither have clear ancestors represented in the fossil record for at least 36 million years before their appearance. If mosasaurs and whales aren't related, why did the planet decide it was time to evolve air-breathing fish-creatures from two disparaging lineages of terrestrial animal only in the Late Cretaceous, and not before or since? There are other marine reptiles with fish-like bodies, such as Ichthyosauromorpha, but they have no fully terrestrial ancestors.
Mosasaurs diverged from Artiodactyla/Ceratops during one of the first two marine catastrophes of the late Cretaceous, and must have lost their horns before true “horn bosses” were a thing. By 53 mya mosasaur had evolved into a fully formed whale. At least one specialized species fed on the coasts of India (then the Himalayas). The shore feeders like himalayacetus and basilosaurus possessed specialized teeth (heterodont), but the deep sea whales probably sported homodont teeth like their mosasaur ancestors. We just don't find many deep sea creatures in the fossil record, extant or extinct, during times when sea life had recently suffered a mass extinction and/or sea levels were lower than today. Therefore our fossil record on mosasaurs and cetaceans is woefully incomplete. Several species of mosasaur have the same number of vertebrae as most mammals. My own assessment at the museum of Georgia Southern University, where a mosasaur and basilosaurus are mounted side by side, revealed no major difference in morphology with the exception of the teeth, temporal fenestra, and number of bones in the jaw. These may all be considered adaptations to three global catastrophes (two of them marine) and/or a shoreline feeding niche.
The similarity between these species is why Basilosaurus is misnamed. Perhaps they had it right the first time.
Aetosaurs, Ankylosaurs, Metatherians, Multituberculata, and Basilosaurus also share a similar type of leaf-shaped teeth not found elsewhere in the fossil record. This suggests a relationship and perhaps even stages in mammalian tooth evolution.
Aetosaur Teeth

Ankylosaur Tooth:


Basilosaur/ Vogtle Whale:


Multituberculata teeth



Mesozoic Mammals:


{kind=link}
The cusps may have aided in the pulling, tearing actions of ornithischians and early mammals. We can suggest the evolutionary relationship of all of the pre-KT animals discussed here by a type of chewing mechanism often used to distinguish herbivorous “dinosaurs” from herbivorous mammals. Aetosaurs and most ornithischians and allotheres chew up and down or back and forth. Placental mammals chew side to side.
The back and forth action coincides with the cheek flanges and the opening of the supraorbital fenestra in larger species. A diapsid skull in mammals must result from adaptation to a larger size and a diet of mainly plants. But part of the placental mammals' ability to survive may have been a rapid change of diet.
Grass.
Grass only arose in the Late Cretaceous. The post-apocalyptic world during and after the three extinction events favored downsizing, economy of structure, and rapid adaptations.
We can see evidence in the way ankylosaurs, ceratopsians, Vintana, and early Xenarthra chewed back and forth instead of side to side, as modern cows do.
Back and forth would be much better for eating leaves off of trees and digging up roots. The muscles attached to those supraorbital fenestras weren't needed as much for a diet of grass, and economy became a means of survival. Sloths and glyptodonts likely kept the bizarre cheek flanges because they lived in a place where vegetation wasn't hit quite so badly by the Late Cretaceous extinctions or the Ktg event that ended it.
'This arrangement allows for the attachment of larger, stronger jaw muscles and enables the jaw to open more widely.” Diapsid, Wikipedia
.”It would grind its food between the teeth in somewhat the same way as a modern rodent, though unlike rodents tritylodontids had a palinal jaw stroke (front-to-back), instead of a propalinal one (back-to-front).”
Tritylodontidae ,Wikipedia
Jutting downward from the sides of the jaw are ‘jugal flanges,’ projections of bone on the homologs of human cheeks. Projections of like these usually serve as muscle attachment sites, and the size of the projection generally reflects the size of the muscle. These facial flanges anchor the masseter muscle, a major chewing muscle that helps close the jaw.
Unlike rodents and similar therians, multituberculates had a palinal jaw stroke (front-to-back), instead of a propalinal (back-to-front) or transverse (side-to-side) one; as a consequence, their jaw musculature and cusp orientation is radically different.[6][10] Palinal jaw strokes are almost entirely absent in modern mammals (with the possible exception of the dugong[12]), but are also present in haramiyidans, argyrolagoideans and tritylodontids, the former historically united with multituberculates on that basis.
Multituberculata, wikipedia
Most ornithiscians also possessed the cheek flanges of Mesozoic mammals and early Xenarthrans, and chewed in a similar fashion:
“Genasauria contains a medial offset of the maxillary dentition (buccal emargination), which is commonly referred to as the ‘ornithischian cheek.'[3] Other characteristics of the ornithischian cheek include “a deep-set position of the tooth rows, away from the sides of the face, a spout-shaped front to the mandibles, and reduction in the size of the opening on the outside of the lower jaw (the external mandibular foramen)."[4] The ornithischian cheek is largely inferred to be evidence for the possession of muscular cheeks that were used for complex chewing behavior and is a fundamentally Genasaurian characteristic.”Genasauria, wikipedia
“It was well adapted for strong, predominantly orthal (up-down) movement for eating rough vegetation.[citation needed] Megatherium possessed the narrowest muzzle of all ground sloths from the Pleistocene. This leads paleobiologists to believe that it was a very selective eater. It had the ability to pick and choose which leaves and twigs it would consume. Megatheriumhad a large narrow prehensile lip that was capable of grabbing and tearing off particular leaves and twigs and other sorts of vegetation.”
Megatherium, Wikipedia
Some ceratopsians were already switching to a more mammalian style of chewing style before the Cretaceous ended:
Leptoceratops wasn’t the only animal with this type of chewing, but Varriale had to look far afield to find other examples. Similar types of tooth wear (and presumably similar types of jaw motions) are found in a handful of small living (some rodents) and extinct (multituberculate and haramiyid) mammals. This dinosaur had independently evolved a chewing style that matched that in those mammals. http://blogs.plos.org/blog/2016/07/07/chew-on-this-mammal-like-mastication-for-the-dinosaur-leptoceratops/
So the cheek flanges may be indicative of larger jaw muscles and are often accompanied by the opening of suprorbital fenestra in synapsids. A need to specialize at the end of the Cretaceous and/or a loss of trees directly following the Ktg forced many mammals to abandon their old diets. For most mammals, the cheek flanges became an expense they just couldn't afford. A side to side choosing action gained favor in ungulates due to a reliance on grass.
Another herbivorous adaptation shared by Multituberculates, ornithischians, and modern herbivorous mammals is the missing area on the front of a jaw. In all of these groups the feature is the same, but we know it to be a hard gum covering in extant mammals. Someone assumed the ornithischian cavity to be a beak long ago, but chances are it was the same as it's “new” relatives. A few have already begun to notice the error of dated assumptions, and propose cheeks and gums for ceratopsians.(7)
And ornithischians show a clear evolutionary progression from using gastroliths to true mastication during the Late cretaceous, whereas “classic” Artiodactyles appear just after 50 million years ago already chewing the cud.
Some may object to this new model of evolution to the difference between the mammalian and ornithischian jaw. Such a discrepancy is often used to point out the difference in the two groups. In truth, the mammalian jaw is a condition resulting from downsizing and a change to a niche environment (possibly a shared stage of a grass-dependent diet). Convergent evolution of a mammalian jaw from ornithischians to mammals is no more of an assumption than convergent evolution of osteoderms and horns. In fact the latter could be argued to be far more complicated processes. We don't even have a good explanation as to how they arose or when. What's more, the new jaw has an explanation: the atmosphere and ecosystem changed radically between the Cretaceous and the Paleogene. Many extant and Paleogene mammals lack the condition.
Most of the mistakes this new model of evolution corrects were made in the late 1800s and early 1900s. We were simply wrong about the difference in synapsids and diapsids, and shouldn't have used them for classification. It's an honest mistake, though. We didn't have as many fossils and we didn't know about continental drift. Continental drift was a fringe theory at the time, as a matter of fact. But there's another assumption which threw us off the correct path many years ago.
The idea that ornithischians were “bird-hipped” as an adaptation to large size caused another mistake we've been dealing with for over a century. No other reptile walks like an ornithischian except mammals.
Saurischians have always been known as far removed from ornithischians, but they've grown farther apart due to new classifications. Their common ancestor has been pushed farther back into the Jurassic. Saurischians walk differently than ornithischians and other reptiles, but it may be due to the fact that most were bipedal or descended from a bipedal ancestor. Varanids, which display a larger size range than any other amniote group in their fossil record, do not adopt an erect stance with larger size.
“Further, paralleling conclusions for the synapsid lineage, these findings suggest that evolution from sprawling to upright posture did not occur in archosaurs as a response to larger size; rather, these archosaurs likely became upright first and larger later. “
Or maybe the ornithischians parallel the synapsids because they arose from synapsids. the above syudy was done to show that an erect walking posture in dinosaurs was not due to size increase. We have to look somewhere else to explain the erect strut of ornithischians, and our paths keep pointing to a synapsid origin. Jurassic and Cretaceous Therapsids and Mammaliaformes walked in the same manner.
So how that all of this escaped our attention for so long, if it's all true?
I think it's because we have been stuck in an aging classification model which did not take Continental Drift into account. The fight to prove the theory of evolution true during the 20th century overshadowed assessment of the overall model itself. Only now have fossils turned up that have allowed me to make these associations. I'm sure others have discovered the same associations, or are discovering them now.
Plus, the period between 66 million years ago and 55 million years ago is scant on fossils. It is a period in which animals had to adapt morphology rapidly in order to survive. The associations between these animals are not immediately recognizable. Different animal groups have different research specialists, who focus on different time periods. They defer to one another when people ask questions, so inquiry turns into a wild goose chase.
I would have never thought closely related synapsids might become diapsids and then independently of one another revert to synapsid form again until I learned the obscure facts I've provided.
It's often said that science progresses only one retirement at a time.
DR. Eugene Anklesloth
Though Dr. Eugene is a fictional character, his explanation of mammalian evolution may be true. All the photos and citations above are real, and the theory makes more sense than the one shown on the Discovery Channel or National Geographic... or even in the universities. Someone needed to take a shot at explaining all of the coincidences the academic majority expects us to believe.
Hi I'm Joe Lyon Layden.
Though Dr. Eugene is a fictional character, his explanation of mammalian evolution may be true. All the photos and citations above are real, and the theory makes more sense than the one shown on the Discovery Channel or National Geographic... or even in the universities. Someone needed to take a shot at explaining all of the coincidences the academic majority expects us to believe.
Hi I'm Joe Lyon Layden.
I began researching our prehistoric past 20 years ago in order to write prehistoric fiction novels. Because mainstream academia's model of evolution contained too many coincidences, inconsistencies, and cases of "special pleading," I knew they must be wrong. I wanted a model of evolution that would stand the test of time, so that people would still enjoy reading my stories a hundred years from now or more.
In the course of my research, I uncovered astounding truths about our planet's history and man's prehistoric past. With the amazing discoveries in archaeology, anthropology, and paleontology over the past few years, I've been able to "put it all together" through deductive reasoning and little-known facts.
I hope to share this with you on this blog and in my novels. The first two stories will be released in 2018 via Radiant Crown Publishing and Immortal Works Press, but you'll get the prequel novella free upon release through the newsletter.
The sign-up button's in the top right column, and I'll also keep you up to date on our ancestor the ankylosaurus with our monthly email! Please check out the photos below, the similarity between the last three skulls in mind-blowing!
A Coincidence of Cheek Flanges?
Ankylosaur


{kind=link}
Ankylosaurus


{kind=link}
Vintana


http://cdn.sci-news.com/images/enlarge/image_2256_2e-Vintana-sertichi.jpg
Giant Sloth Skull


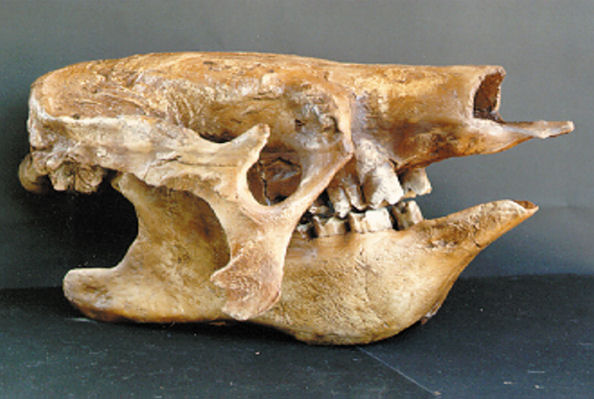{kind=link}
http://fossilexpeditions.com/gifarma1.jpg
Glyptodont Skull:


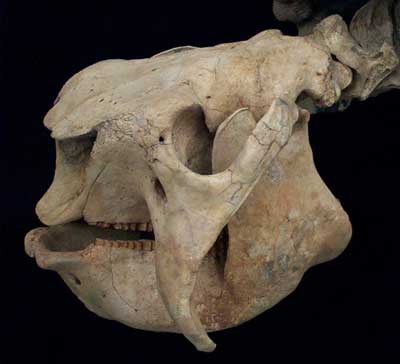{kind=link}
And the possible mother of them all, appearing 267 million years ago in the Therapsid lineage just before the dawn of Aetosaurs and Trittylodontids:


{kind=link}
Estemmenosuchus has everything evolution needs to make mammals and ornithischians of all shapes and styles, from osteoderms to horn bosses to cheek flanges. Could this be the Therapsid who started it all?
Ask your professor how these skulls are more different from one another than the last two are from that of a shrew:
Ankylosaur:

http://www.dinocasts.com/images/products/gastonia_web(2).jpg
Giant Sloth (Xenarthra)

http://fossilexpeditions.com/gifarma1.jpg
Pampathere (Xenarthra):

https://en.wikipedia.org/wiki/Pampatheriidae#/media/File:Holmesina_3_Clean.png
(1)"They are reliable markers for inferring evolutionary history, the researchers explain, because their integration into the genome is random—making it highly unlikely for the same element to integrate independently into a conserved region of the genome (called an orthologous position) in two different species. In addition to finding “significant support” for the previously identified divisions, they offer strong support for placing Xenarthra—armadillos and their kin—at the base of the placental tree."
http://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.0040111
The authors, using human chromosomes as probes to study three Xenarthra species, found that one of them, the two-toed sloth Choloepus hoffmannii (2n = 50), has a karyotype strikingly similar to the proposed 2n = 48 ancestral Eutherian complement. This observation, besides reinforcing the hypothesized karyotype, suggests that Xenarthra may be at the root of the Eutherian tree.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1513266/
Extinct glyptodonts really were gigantic armadillos, ancient DNA shows
https://phys.org/news/2016-02-extinct-glyptodonts-gigantic-armadillos-ancient.html
http://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.0040111
The authors, using human chromosomes as probes to study three Xenarthra species, found that one of them, the two-toed sloth Choloepus hoffmannii (2n = 50), has a karyotype strikingly similar to the proposed 2n = 48 ancestral Eutherian complement. This observation, besides reinforcing the hypothesized karyotype, suggests that Xenarthra may be at the root of the Eutherian tree.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1513266/
Extinct glyptodonts really were gigantic armadillos, ancient DNA shows
https://phys.org/news/2016-02-extinct-glyptodonts-gigantic-armadillos-ancient.html
(2) Genomic evidence reveals a radiation of placental mammals uninterrupted by the KPg boundary https://pdfs.semanticscholar.org/dbc3/a416c3c24eea1d2455b65eb479c20a6b3ea5.pdf
http://blogs.plos.org/paleocomm/2016/07/06/chew-on-this-mammal-like-mastication-for-the-dinosaur-leptoceratops/
(5)
No comments:
Post a Comment